


华西耳鼻喉学术前沿速递——文献精读(第52期)
精读分享│【Immunity】转录因子NF-kB在对Toll样受体4信号的初级反应中协调核小体重塑
英文题目:The transcription factor NF-kB orchestrates nucleosome remodeling during the primary response to Toll-like receptor 4 signaling
中文题目:转录因子NF-kB在对Toll样受体4信号的初级反应中协调核小体重塑
期刊:Immunity(IF:25.5)
单位:美国加利福尼亚大学洛杉矶分校
发表时间:2024年2月
摘要:
在巨噬细胞中,数百个潜在增强子和多个启动子在可诱导核小体重塑的过程中塑造对Toll样受体4(TLR4)信号的转录反应。本研究旨在确定那些促进TLR诱导重塑的转录因子。基于ATAC-seq和单细胞ATAC-seq的分析策略,本研究富集了最可能发生重塑的基因组区域,并发现转录因子核因子kB(NF-kB)结合到标记初级反应期间重塑的所有高可信峰上,这一反应可以由TLR4配体脂质A激活。NF-kB亚基RelA和c-Rel的缺失导致在高可信ATAC-seq峰上的重塑丧失,而对NF-kB结合位点进行CRISPR-Cas9突变也损害了重塑过程。特定区域的重塑选择性由其他诱导因子的协同作用所决定,包括IRF3和MAP激酶诱导的因子。因此,NF-kB在TLR4激活的转录因子中具有独特性,既能广泛促进诱导性核小体重塑,又具有激活开放染色质中的已定位增强子和启动子的能力。
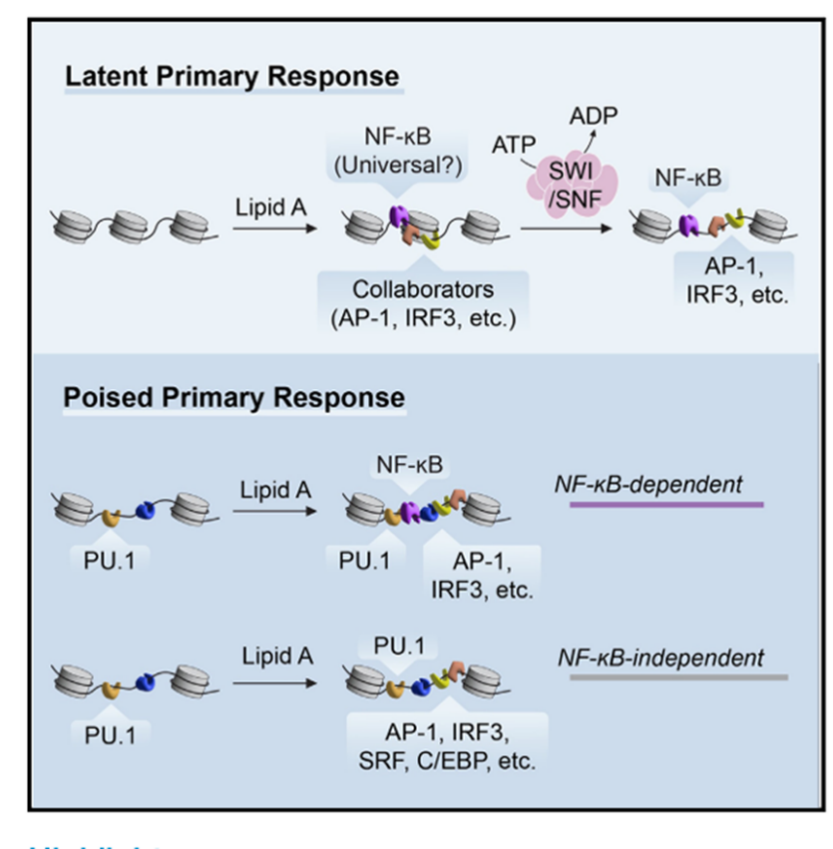
研究背景:
先天免疫细胞需要选择性地激活数百个基因,以对多种刺激做出特定的反应。这种反应的选择性由识别刺激的传感器、传感器激活的下游信号通路,以及通路激活的转录和染色质调节因子共同决定。
核因子κB(NF-kB)是一类通过翻译后修饰机制被激活的转录因子,它们结合在特定的基因调控区域以激活转录。
TLR4激活引发的信号通路会通过调控NF-κB等转录因子,启动数百个基因的表达,从而调节细胞的炎症反应和抗病毒反应。
刺激响应性的核小体重塑通常由ATP依赖的重塑复合体催化的,包括SWI/SNF复合体,这些复合体可以促进核小体的驱逐、滑动或构象转变,从而调控染色质结构和基因表达。
一个关键的未解决的问题是,TLR诱导的染色质重塑具体由哪些转录因子发挥作用。该研究重点探索了NF-κB在核小体重塑过程中的作用,特别是在TLR4激活后,NF-κB如何与其他转录因子(如IRF3)协同作用以调节染色质结构。
实验模型及方法:
实验模型:
小鼠:C57BL/6J、Irf3-/-
细胞系:J2-transformed WT BMDM lines、NF-kB motif mutant lines、Rela-/-Rel-/-mutant lines、J2-transformed Nfkb1-/- BMDM lines、Nfkb1-/- Nfkb2-/- mutant lines、Ccl5 promoter mutant lines、Ccl5 upstream region mutant lines
方法:
Stimulation、ATAC-seq、scATAC-seq、RNA-seq、ChIP-seq、CRISPR-Cas9 Ribonucleoprotein(cRNP) genome editing、qRT-PCR、4Cassay、Promoter capture Hi-C assay
研究思路:

研究结果:
1.ATAC-seq分析揭示了广泛的TLR4诱导峰值
方法:
对小鼠骨髓来源的巨噬细胞(BMDMs)进行了ATAC-seq分析,这些细胞被TLR4配体脂多糖A刺激了0、30、60或120分钟。
结果:
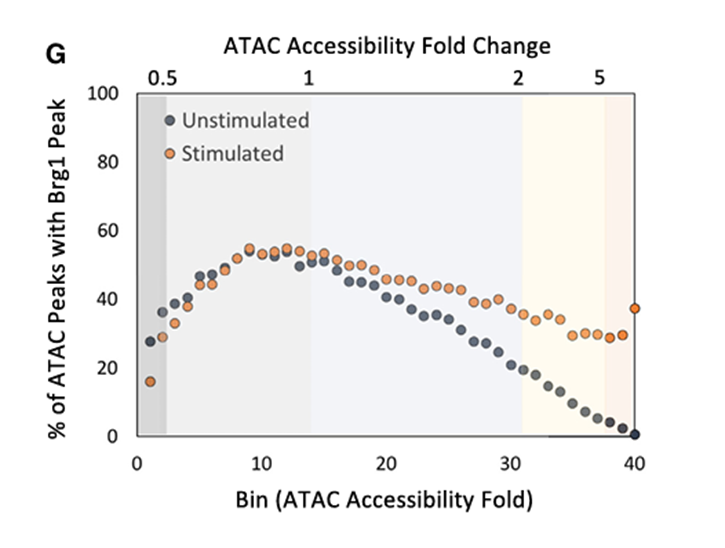
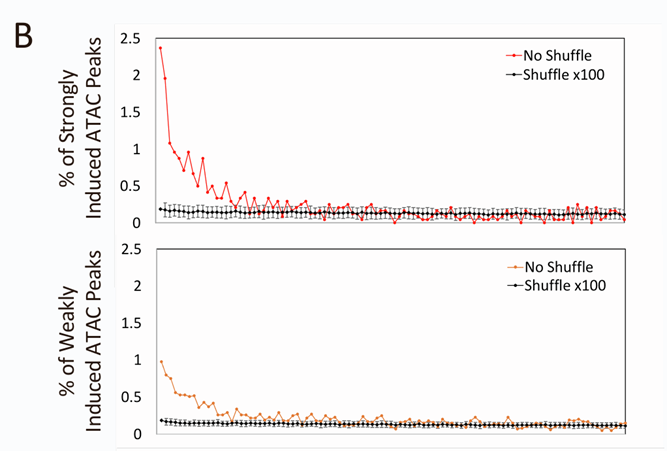
多数峰值(72.8%)的变化幅度小于2倍,2.7%的峰值表现出超过2倍的变化,而19.4%的峰值被诱导至2到5倍之间,其中5.1%(5,178个峰值)诱导超过5倍。大多数峰值位于内含子和基因间区域。
尽管大多数诱导超过2倍的峰值在统计上是显著的(p<0.05),但其中有很大一部分在未经刺激和刺激的细胞中均可观察到,随着每个区间内平均激活倍数的增加,未经刺激细胞中的峰值的比例会逐渐下降。
研究者特别关注ATAC-seq信号增强超过5倍的峰值,并提高了统计标准以进一步强调最有可能经历核小体重塑的区域。最强烈且一致诱导的ATAC-seq信号来表征核小体重塑。

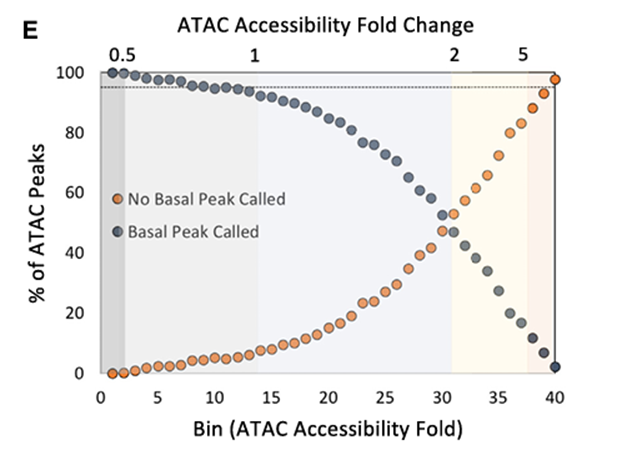
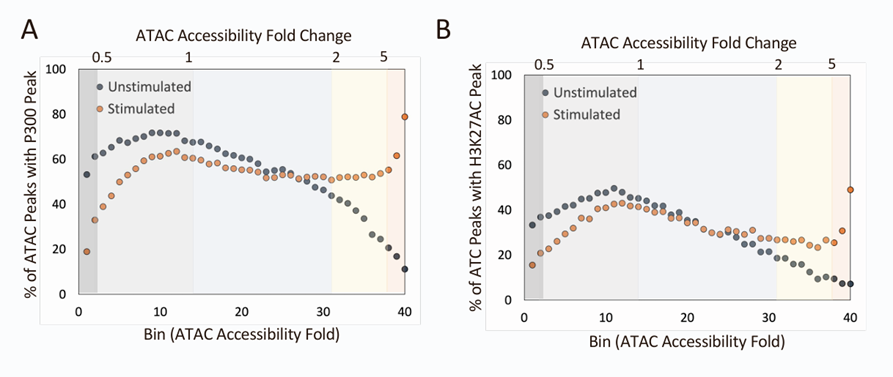
2.ATAC-seq信号的诱导与可诱导的BRG、p300和H3K27Ac ChIP-seq峰值相一致
方法:
用SWI/SNF复合体的Brg1催化亚基进行ChIP-seq。
结果:
在未刺激的骨髓来源的巨噬细胞(BMDMs)中,小幅度诱导的ATAC-seq峰值区域中检测到Brg1峰值的比例为20%至50%,并且在刺激后这些比例几乎没有变化。相比之下,在40号分箱中,未刺激的细胞中Brg1 ChIP-seq峰值仅在0.8%的ATAC-seq峰值区域被观察到,但在刺激后这一比例上升到37%。
针对p300(一种转录共激活因子)和组蛋H3K27ac(一种与活跃转录相关的组蛋白修饰),也得到了类似的结果。
这些结果支持了这样一个假设:相比于表现出较弱ATAC-seq诱导峰值的区域,显示出最强ATAC-seq诱导峰值的区域更有可能经历TLR4诱导的Brg1招募和核小体重塑。
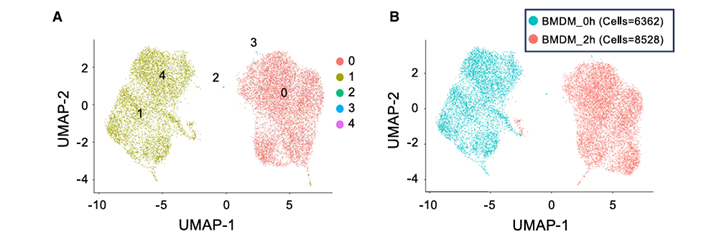
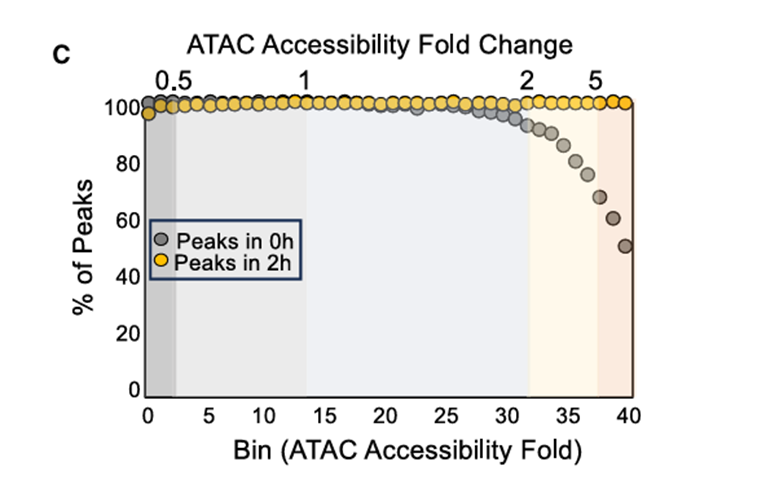
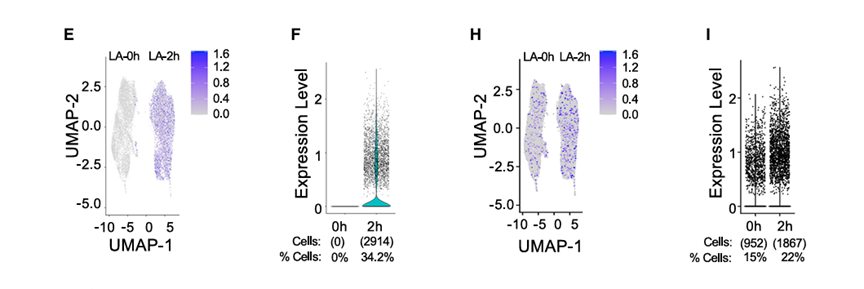
3.单细胞ATAC-seq确定强烈诱导峰值和弱诱导峰值之间的差异
方法:
对使用脂多糖A刺激后的BMDMs进行了单细胞ATAC-seq(scATAC-seq),分别在0小时和2小时取样。
结果:
1)只在40个分箱中的最后一个分箱中观察到未刺激和刺激细胞之间的巨大差异
scATAC-seq研究结果与假设相符,即ATACseq信号的适度、统计学上显著增加是由于染色质或转录因子结合的适度变化引起的,这并不代表真正的核小体重塑。
4.在启动子和基因间区域的可诱导ATAC-seq峰的特征
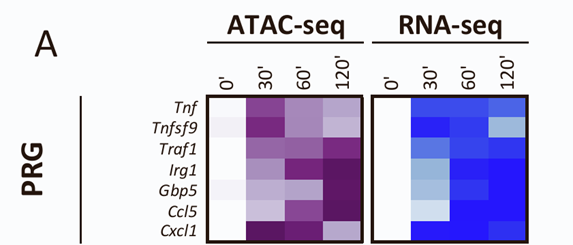
方法:回到批量分析、RNA-seq
结果:之前的研究集中在TLR4信号传导有效诱导的132个主要反应基因的启动子上,结果发现只有三个基因的启动子(Ccl5、Gbp5和Irg1)显示出强烈的ATAC-seq诱导,结果证实,启动子处的强ATAC-seq信号诱导是非常罕见的。
方法:通过计算每个TLR4诱导的ATAC-seq基因间峰值到最近的、表现出至少10倍转录诱导的基因的距离,来分析基因间区域。。
结果:强诱导的ATAC-seq峰(>5倍)在强诱导基因附近显著富集;弱诱导的ATAC-seq峰在较小程度上也呈现过度富集。
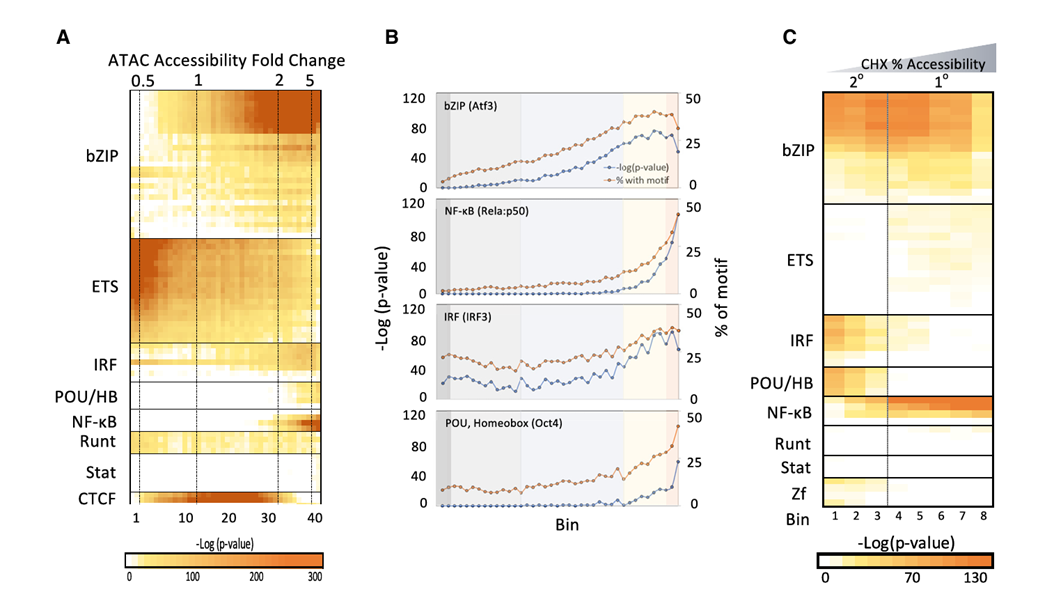
5.在主要响应的ATAC-seq峰值中NF-kB基序富集的独特轮廓
方法:基序分析
结果:在包含强诱导的ATAC-seq峰的分箱中,bZIP、ETS、IRF、NF-kB和POU/HB的基序富集程度最高。与弱诱导的分箱相比,NF-kB基序与最强诱导的ATAC-seq峰分箱关系最密切。
方法:在CHX(区别初级、次级反应)存在的情况下,对BMDM进行ATAC-seq
结果:NF-kB基序在表现出最强CHX抗性(初级响应)的峰中富集最显著。
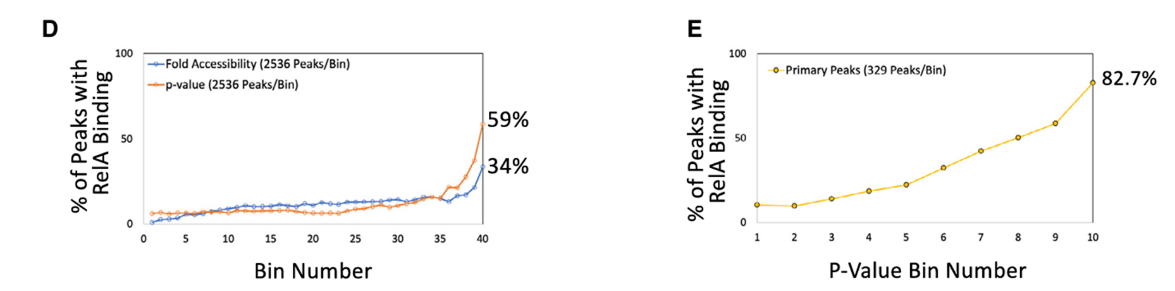
6.NF-kB广泛结合于高置信度的初级反应ATAC-seq峰
方法:检查NF-kB家族成员RelA的ChIP-seq数据
结果:在基于倍数诱导划分的40个ATAC-seq峰的分箱时,RelA结合最常出现在最高倍数诱导的分箱(分箱40),其中34%的峰与RelA结合。当根据ATAC-seq诱导的p值创建40个分箱时,最高置信度分箱中59%的峰表现出RelA结合。
方法:专注于3289个表现出强诱导(>5倍)的初级响应峰,并根据它们的p值将它们分成10个区间。
结果:在最高置信区间(分箱10)中,82.7%(272/329)的峰与RelA ChIP-seq峰重合。
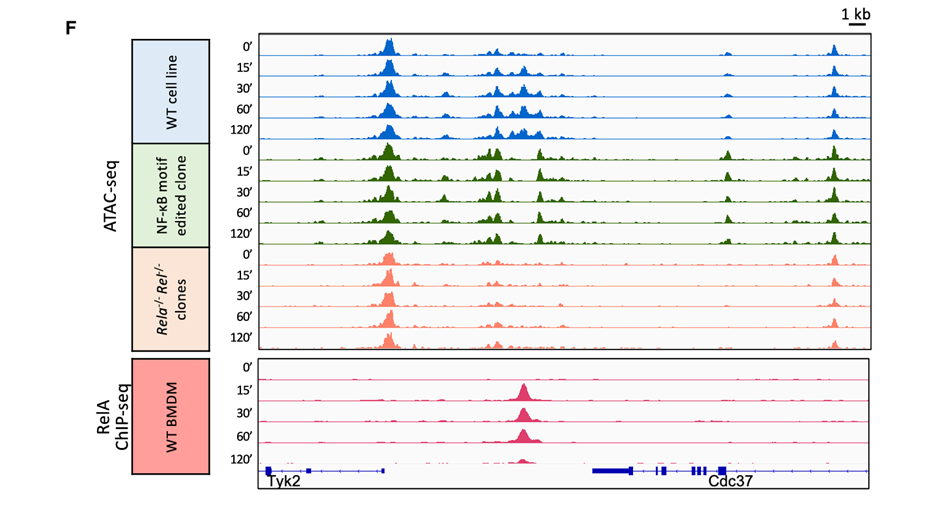
7.NF-kB基序以及Rela和Rel基因在ATAC-seq激活中具有重要作用
方法:对Tyk2和Cdc37基因之间的一个强诱导ATAC-seq峰下的两个NF-κB基序进行了置换突变(使用一个J2病毒转化的小鼠巨噬细胞系)。
结果:在突变细胞系中,该基因间区域的可诱导ATAC-seq信号选择性缺失(轨道6-10),表明NF-κB结合对信号对诱导至关重要。
方法:使用CRISPR-Cas9编辑构建了突变的巨噬细胞系,在这些细胞系中引入了Rela和Rel(c-Rel)基因的纯合缺失(Rela⁻/⁻Rel⁻/⁻细胞系)。
结果:在两个独立的Rela⁻/⁻Rel⁻/⁻克隆细胞系中,Tyk2/Cdc37区域的可诱导ATAC-seq信号完全消失了(轨道11–15)。这一结果进一步证明了NF-κB在该区域的核小体重塑中不可或缺的作用。
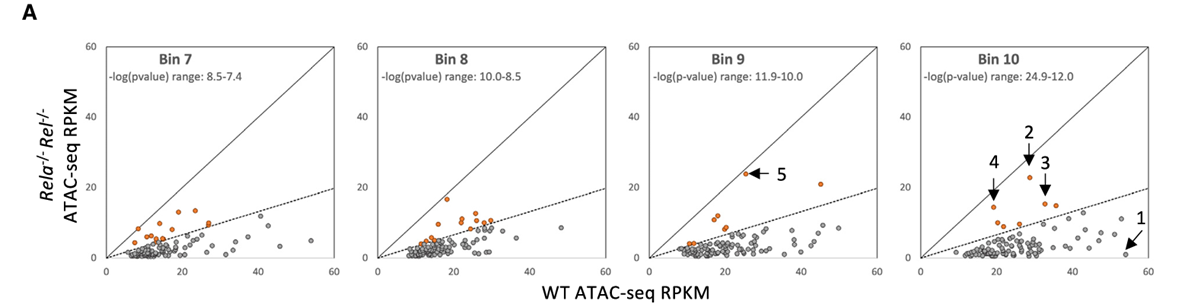
8.RelA和c-Rel在高置信度ATAC-seq峰上的广泛影响
方法:分析Rela⁻/⁻Rel⁻/⁻克隆细胞在刺激后获得的ATAC-seq数据。研究了在野生型细胞系中诱导超过5倍的1001个初级反应峰值,并根据诱导的统计显著性(p值)将这些峰分为10个区间。
结果:这些突变对ATAC-seq信号的影响(RPKM),主要集中在统计显著性最强的四个区间。在每个区间中,大约90%的峰值在Rela⁻/⁻Rel⁻/⁻克隆细胞中显著减少(强度不到野生型的33%)。受影响较小的峰值主要属于以下两种情况:(1)部分次级反应中不依赖NF-κB(2)存在转录读穿现象。
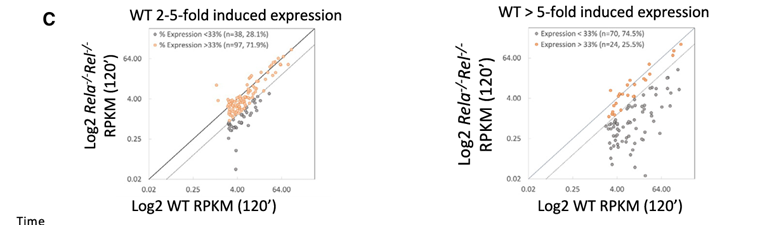
方法:对野生型(WT)和Rela⁻/⁻Rel⁻/⁻突变细胞系进行RNA-seq分析,比较基因转录水平的变化。
结果:在诱导超过5倍、且RPKM达到3的基因中,突变使74.5%的基因(94个基因中的70个)的mRNA水平相比于野生型显著下降。相比之下,在诱导水平为2至5倍的基因中,只有28.1%的基因(135个基因中的38个)的转录水平显著下降(图4C,左)。
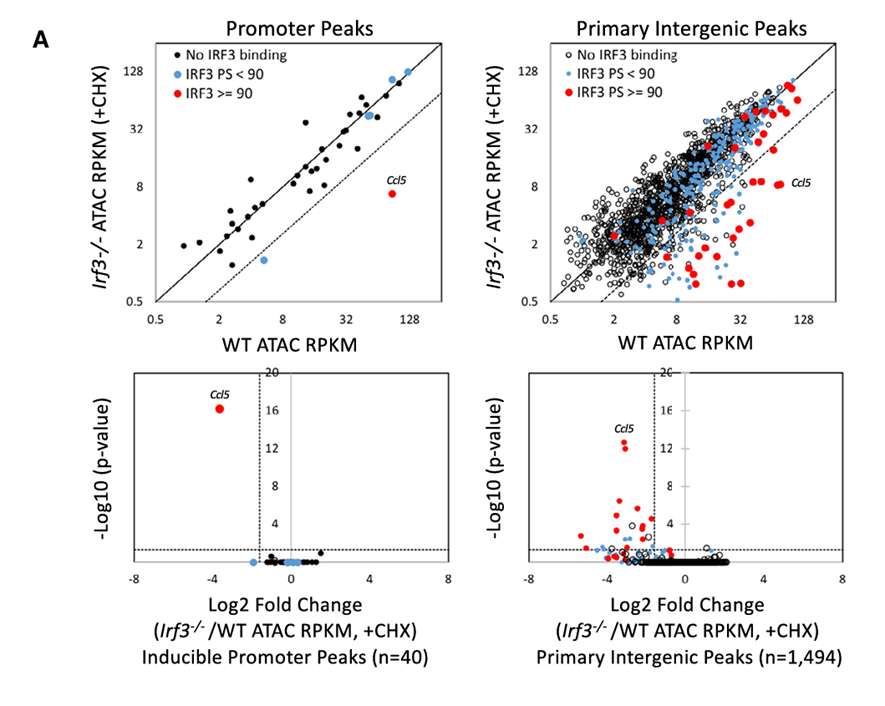
9.IRF3在少量ATAC-seq峰的诱导重塑中的关键作用
方法:对Irf3⁻/⁻ BMDMs进行ATAC-seq和ChIP-seq,研究IRF3在基因间区的染色质重塑作用。
结果:Ccl5启动子在其IRF3需求上表现出独特性(图5A,左)。
方法:将诱导性的基因间区ATAC-seq峰在野生型(WT)和Irf3⁻/⁻BMDMs(在存在CHX的情况下刺激以专注于初级反应)中进行了比较。
结果:在散点图中(图5A,右上),只有107个诱导性基因间位点表现出对IRF3的强依赖性;其中19个位点在ChIP-seq分析中表现出强烈的IRF3结合(图5A中的红点)。在火山图中(图5A,右下),仅有22个基因间区的IRF3依赖性初级反应ATAC-seq峰具有统计学显著性,其中有13个(59%)表现出强烈的IRF3结合。
在初步的ATAC-seq分析中,IRF基序在初级反应中并未富集。然而,对IRF3依赖性的ATAC-seq峰进行的基序分析显示,IRF基序明显富集。因此,最初分析中未检测到IRF基的富集,是因为IRF3只在一小部分可诱导的峰中发挥重要作用。
10.NF-κB和IRF3都在Ccl5基因位点的染色质重塑中起到了关键的作用
方法:为了进一步探究NF-κB与IRF3之间的协作,研究者聚焦于Ccl5位点,该位点包含两个对IRF3高度依赖的高置信度ATAC-seq峰,分别位于Ccl5启动子以及Ccl5转录起始位点(TSS)上游10kb处(IRF3依赖性峰用红色和蓝色阴影标记)。
结果:在Rela⁻/⁻Rel⁻/⁻突变细胞系中,所有的可诱导ATAC-seq峰都消失了,Ccl5 mRNA的诱导也大幅减少。在Irf3⁻/⁻BMDMs中也同样如此。
因此,IRF3和NF-κB都对这两个IRF3依赖性位点的可诱导ATAC-seq是必需的,证实了这两个因子可以协作以促进染色质重塑。

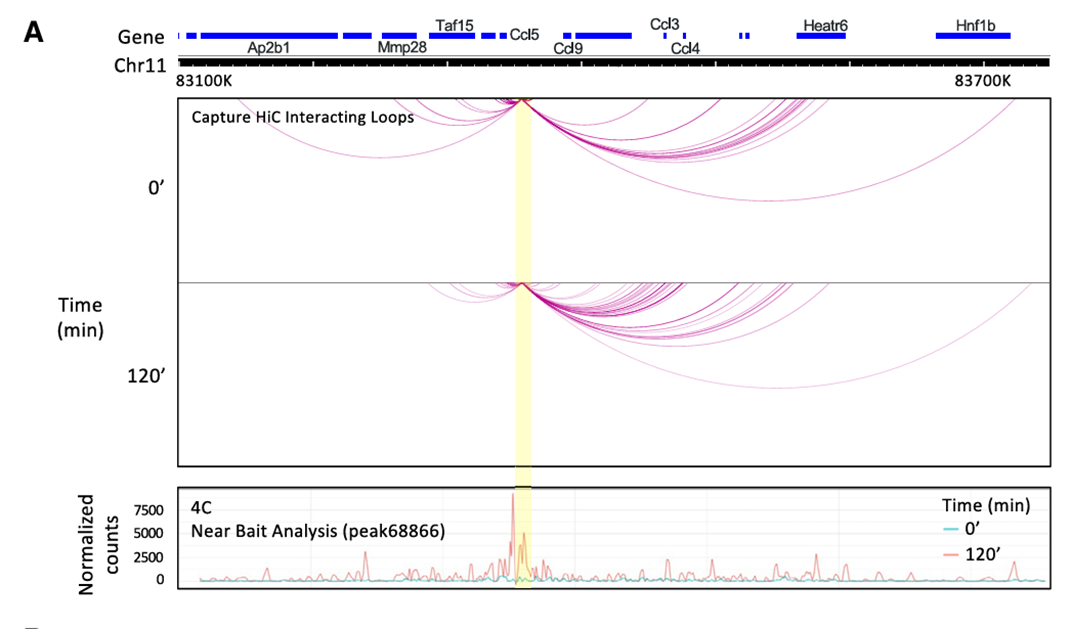
11.Ccl5位点的可诱导ATAC-seq峰区域支持DNA环状结构和其功能
方法:为了进一步研究Ccl5基因间区位点,进行了环状染色体构象捕获(4C)和捕获Hi-C实验。将对应于基因间IRF3依赖性ATAC-seq峰的DNA区域作为目标。
结果:在未刺激的BMDMs中,目标区域与远端位点之间的相互作用数量有限,而在刺激后相互作用显著增加。最广泛的、可诱导的相互作用发生在Ccl3和Ccl4基因附近。
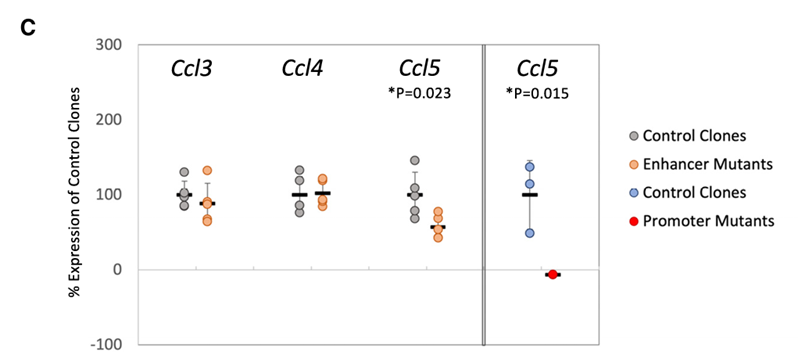
方法:使用CRISPR敲除了对应IRF3依赖性ATAC-seq峰的基因组区域。作为对照,单独删除了Ccl5启动子。
结果:在三个缺失Ccl5启动子的独立克隆中,通过RT-qPCR分析发现Ccl5转录被完全抑制。敲除基因间区域也减少了Ccl5转录,但幅度较小。而敲除该区域对Ccl3或Ccl4转录没有影响。

汇报人:吴婷婷
导师:任建君
审核:夏晓旭、冯兰、任建君
