


精读分享│【Nature Medicine】:吉西他滨联合顺铂治疗后鼻咽癌的肿瘤免疫微环境
英文题目:The tumor immune microenvironment of nasopharyngeal carcinoma after gemcitabine plus cisplatin treatment
中文题目:吉西他滨联合顺铂治疗后鼻咽癌的肿瘤免疫微环境
期刊:Nature Medicine(IF: 58.7)
单位:中山大学肿瘤防治中心
发表时间:2023年6月
PMID: 37280275
DOI: 10.1038/s41591-023-02369-6



摘要:
吉西他滨联合顺铂(GP)化疗是鼻咽癌(NPC)的标准治疗方案。然而,其临床疗效的机制尚不清楚。该研究对比了15对匹配的、未接受过治疗的和接受GP化疗后的NPC样本,进行了单细胞RNA测序与T细胞和B细胞受体测序,发现GP化疗激活了由先天样B细胞(ILB)主导的抗肿瘤免疫反应。化疗诱导的DNA片段激活了STING-I型干扰素依赖性通路,增加了癌细胞中主要组织相容性复合体Ⅰ类分子的表达,同时通过Toll样受体9信号通路诱导ILB。ILB通过ICOSL-ICOS轴进一步扩增滤泡辅助性T细胞(Tfh)和辅助性1型T细胞(Th1),从而在化疗后增强生发中心缺乏的三级淋巴器官样结构中的细胞毒性T细胞。在一项针对接受GP化疗的NPC患者的3期临床试验中(NCT01872962, n = 139),ILB的频率与总生存期和无病生存期呈正相关。此外,它还可以作为预测GP 和免疫疗法联合治疗在NPC 患者良好结局的潜在标志物 (n = 380)。总的来说,该研究为GP化疗后肿瘤免疫微环境提供了高分辨率图谱,揭示了以B细胞为中心的抗肿瘤免疫的作用。本研究确定并验证了ILB作为NPC基于GP治疗的潜在生物标志物,有望改善患者管理。
研究内容思维导图:

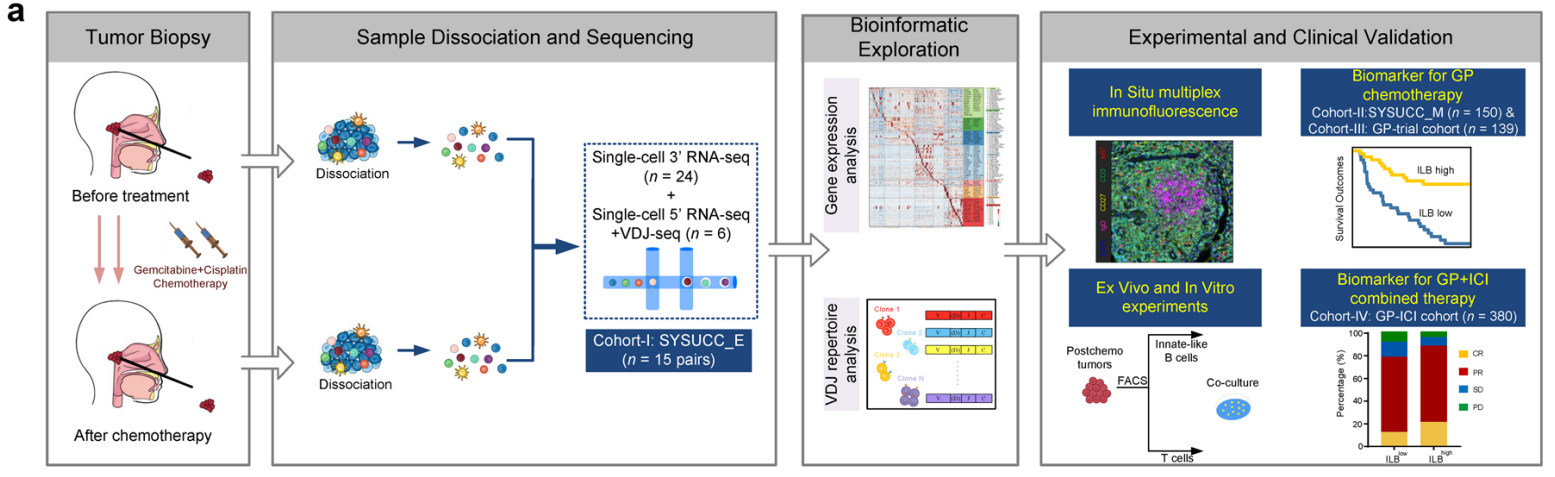
主要技术路线图:
研究简介:
1.背景与研究目的:
化疗的疗效不仅依赖于直接的细胞毒性作用,还依赖于激活抗肿瘤免疫反应。然而,目前尚不完全理解临床有效的化疗如何驱动肿瘤微环境中的信号传导和成分变化,以引发最佳的抗肿瘤反应。现有研究主要关注于化疗诱导的肿瘤抗原暴露或濒死细胞释放的危险信号(即免疫原性细胞死亡)。然而,仅仅靠抗原或危险信号的释放不足以克服肿瘤免疫逃逸,长期抗原暴露甚至可能导致免疫耐受。
2.挑战与知识空白:
成功的化疗引发的抗肿瘤反应所需的免疫学变化尚未完全被揭示。比如,死亡细胞的残骸需要由功能性树突状细胞(DCs)处理和呈递,以引发抗原特异性T细胞反应。但在肿瘤微环境压力下,DCs常常功能受损或表现为免疫耐受。此外,B细胞在肿瘤免疫微环境中作为抗原呈递细胞(APCs)启动滤泡辅助性T细胞(Tfh)反应的作用,除其抗体分泌功能外,仍未得到充分探索。
3.研究意义:
目前,关于B细胞是否能够触发抗肿瘤T细胞反应,以及化疗如何改变B细胞的环境信号和功能表型的数据较少。吉西他滨(GEM)联合顺铂(DDP)化疗方案(即GP方案)在治疗多种癌症中广泛使用,尤其是在鼻咽癌(NPC)中显示出显著疗效。鼻咽癌作为一种具有高度转移性的侵袭性头颈癌,为研究化疗如何重塑肿瘤免疫微环境的提供了理想的模型。
4.研究方法与主要发现:
①研究全面表征了鼻咽癌患者在GP化疗后的肿瘤免疫微环境(TIME)的演变模式,并识别出先天性样B细胞(ILBs)在抗肿瘤免疫反应中占主导地位。②通过多学科方法,研究确定了ILBs的诱导机制及其与T细胞的功能性相互作用,显示出增强的抗肿瘤潜力,同时伴随着肿瘤细胞中主要组织相容性复合体(MHC)I类分子的表达增加。③ILBs被提出作为鼻咽癌GP化疗的潜在生物标志物。
主要结果:
1、 GP化疗重塑NPC的肿瘤免疫微环境(TIME)
该研究团队选择了15名NPC患者,收集了中山大学肿瘤防治中心(SYSUCC)GP化疗前后的30个配对肿瘤样本(Cohort_1, SYSUCC_E cohort)。通过10X Genomics Chromium微滴单细胞RNA测序(scRNA-seq)技术,共获得216,893个单细胞转录组,并对其中6个配对样本进行了单细胞T细胞和B细胞受体测序(scTCR-seq和scBCR-seq)(图1a)。研究将细胞分为七个主要群体,并进一步细分为86个亚群。结果显示,化疗前后T细胞和B细胞在NPC样本中的免疫成分中占比超过80%,特别是非浆细胞型B细胞在化疗后显著增加,而T细胞和髓系细胞的频率并未显著变化(图1b)。单细胞轨迹分析表明,化疗促使癌细胞从高增殖状态向两条不同的轨迹分化,其中主要轨迹(Trajectory I)与抗原呈递分子的显著表达相关(图1c)。此外,化疗后MHC I类分子和抗原呈递基因的表达增加,特别是在对化疗有响应的肿瘤中,这一结果通过流式细胞术分析得到了进一步验证(图1d)。研究还发现Trajectory I特征的癌细胞与患者的良好预后显著相关,提示化疗引发的抗原呈递能力增强可能是预后改善的重要因素(图1e)。
研究团队进一步探讨了GP化疗如何增强NPC细胞中MHC I类分子和抗原呈递基因的表达。单细胞数据集显示,化疗后的NPC样本中,I型干扰素(IFN-I)信号通路以及对IFN-I信号的反应显著增强,伴随着MHC I类分子表达的增加(图1f)。在图1d中MHC I类分子表达增加的样本中,化疗后肿瘤中的干扰素-β(IFNβ)水平比未经治疗的配对样本高出三倍(图1g)。研究还观察到下游信号分子STAT1的显著激活。GP化疗产生了协同效应,进一步促进了IFN信号相关的MHC I类分子和抗原呈递基因的表达(图1h, i)。此外,化疗后NPC样本中观察到DNA传感器STING的显著增加及其激活的增强,尤其是在MHC I类分子表达增加的情况下(图1k)。GP化疗在NPC细胞中协同激活了STING–TBK1–IRF3信号通路(图1l)。研究团队使用CRISPR–Cas9系统敲除HK1细胞中的STING基因,发现这一干预有效阻止了化疗诱导的IFNβ产生、STAT1的激活以及MHC I类分子和抗原呈递基因的表达增加(图1m)。类似的结果在STING敲低的HONE1细胞中也得到了验证。进一步实验显示,在STING敲除的HK1细胞中过表达STING可以恢复GP治疗诱导的IFNβ产生(图1n)。因此,研究表明GP化疗可以通过STING依赖的IFN-I通路增强NPC细胞中MHC I类分子相关的抗原呈递机制。
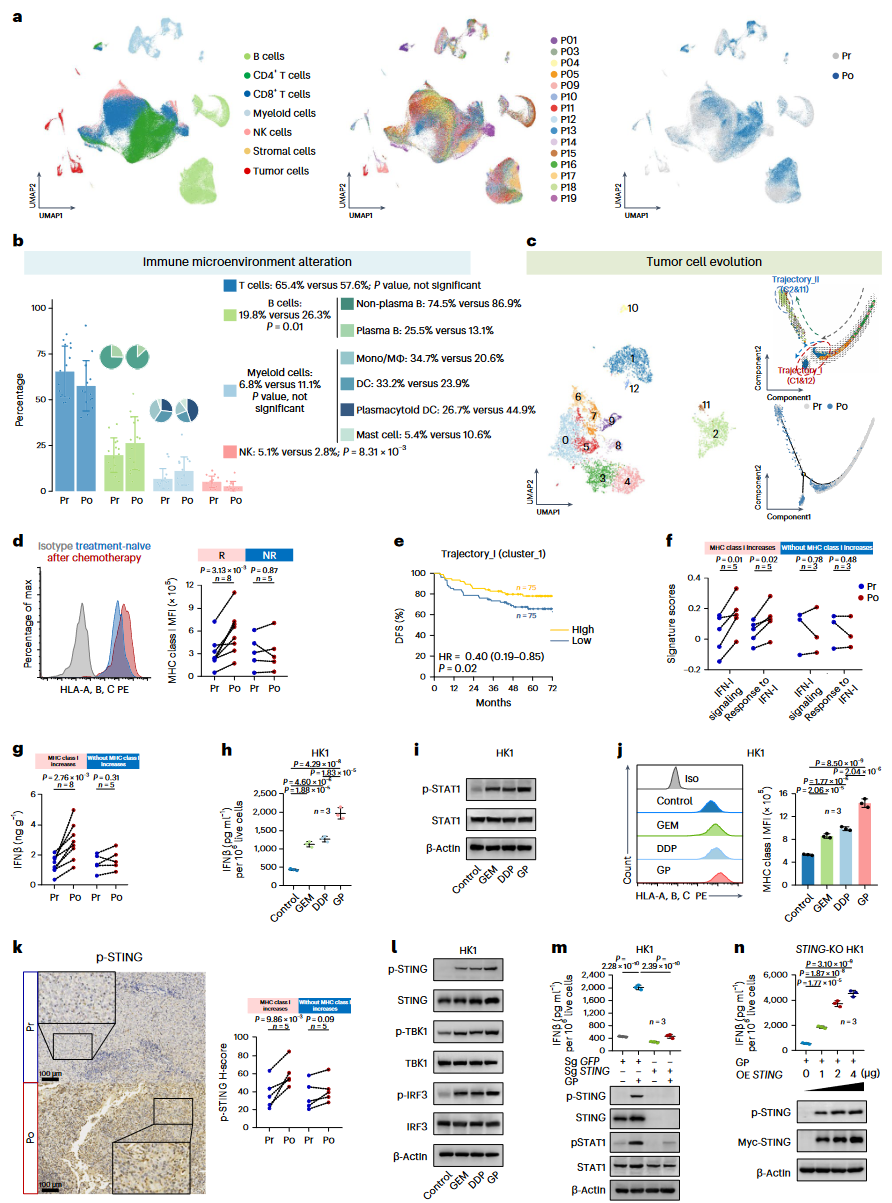
图1:GP化疗重塑NPC的肿瘤免疫微环境
2、GP化疗触发Th1样和Tfh诱导的CTL反应
研究团队进一步探索了GP化疗后NPC中免疫细胞亚群的变化。研究发现,在CD8+ T细胞中,化疗后出现了显著的群体变化,具体表现为初始型CD8+ T细胞(Cluster_0)、活化型CD8+ T细胞(Cluster_2)和两个细胞毒性T淋巴细胞(CTL)亚群(Cluster_3和10)显著增加,而耗竭T细胞(Cluster_5)的比例下降(图2a)。此外,化疗后CD8+ T细胞的TCR特征发生了变化,每个CD8+ T细胞群体中都出现了新的克隆,其中新克隆在耗竭T细胞(Cluster_5)中占比超过70%(图2a)。这一现象表明,化疗后新的T细胞克隆可能经历早期活化并随后进入耗竭状态。与这一观察一致,化疗后的肿瘤裂解物比化疗前更有效地触发自体CD8+ T细胞产生干扰素-γ(IFNγ)(图2b)。
在CD4+ T细胞中,TCR谱的变化更加显著,化疗后14个CD4+ T细胞亚群中的新克隆比例超过90%(图2c)。研究发现,Tfh细胞(Cluster_6)和Th1样T细胞(Cluster_7)的比例在化疗后增加,这些细胞已被报道可维持CTL反应(图2c)。研究还发现,从化疗后的NPC肿瘤中分离的CD4+ T细胞能够有效促进自体CD8+ T细胞的细胞毒性功能,这一过程在中和白细胞介素-21(IL-21)或阻断干扰素γ受体-1(IFNγR1)时显著受损(图2d)。在临床样本中,Th1样和Tfh细胞与颗粒酶B(GZMB)+ CD8+ T细胞呈正相关(图2e)。生存分析显示,TH1样和Tfh细胞与良好的预后显著相关,这一发现通过单变量和多变量生存分析得到了验证(图2f)。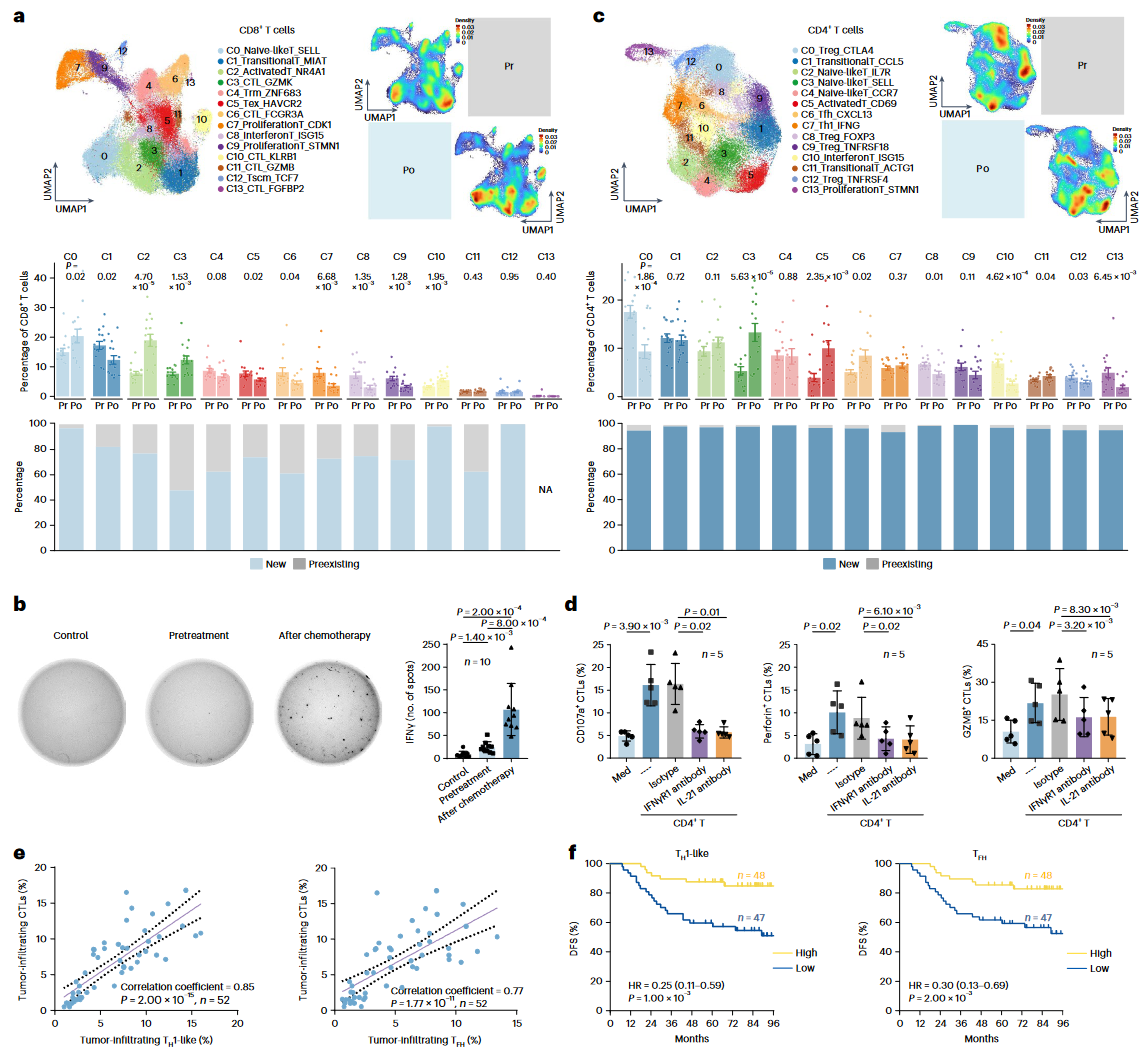
图2:GP化疗触发Th1样和Tfh诱导的CTL反应
为了探讨NPC化疗后的肿瘤免疫微环境是如何促进CD4+ T细胞的效应反应,研究团队发现髓系抗原呈递细胞(APCs),如经典树突状细胞(DCs)和肿瘤相关巨噬细胞(TAMs)的频率在化疗后并未显著增加,甚至有所下降(图3a)。此外,从肿瘤样本中分离出的树突状细胞(tDCs)更倾向于诱导Treg细胞,而非激活Th1和Tfh细胞(图3b)。有趣的是,化疗后B细胞与CD4+ Th细胞的相互作用评分高于髓系APCs,尤其是与Tfh和Th1样细胞的相互作用(图3d)。
通过分析B细胞的单细胞数据,发现有两个显著增加的B细胞亚群(Cluster_5和Cluster_8),其在未经治疗的肿瘤中占比为7.3%,但在化疗后增加至18.5%(图3e)。这两个B细胞亚群表现出高水平的CD25、NR4A1和NR4A2,提示其处于激活状态(图3f)。与记忆B细胞和浆细胞相比,这些B细胞群体显示出较低的VH区域体细胞高突变(SHM)和较高的BCR多样性,并且IGHD和IGHM的表达增加,表明其处于原始阶段(图3g-h)。
已经证明ICOSL信号通路驱动生发中心(GCs)的Tfh分化。ICOSL信号在ILBs中显著升高,并在化疗后进一步增加,这与Th1和Tfh细胞中的ICOS表达增加相对应(图3k)。ILBs与Tfh和Th1样细胞之间的ICOSL依赖性相互作用评分也很高(图3l),表明ILBs在化疗后的NPC微环境中可能对Th细胞的调控起到重要作用。
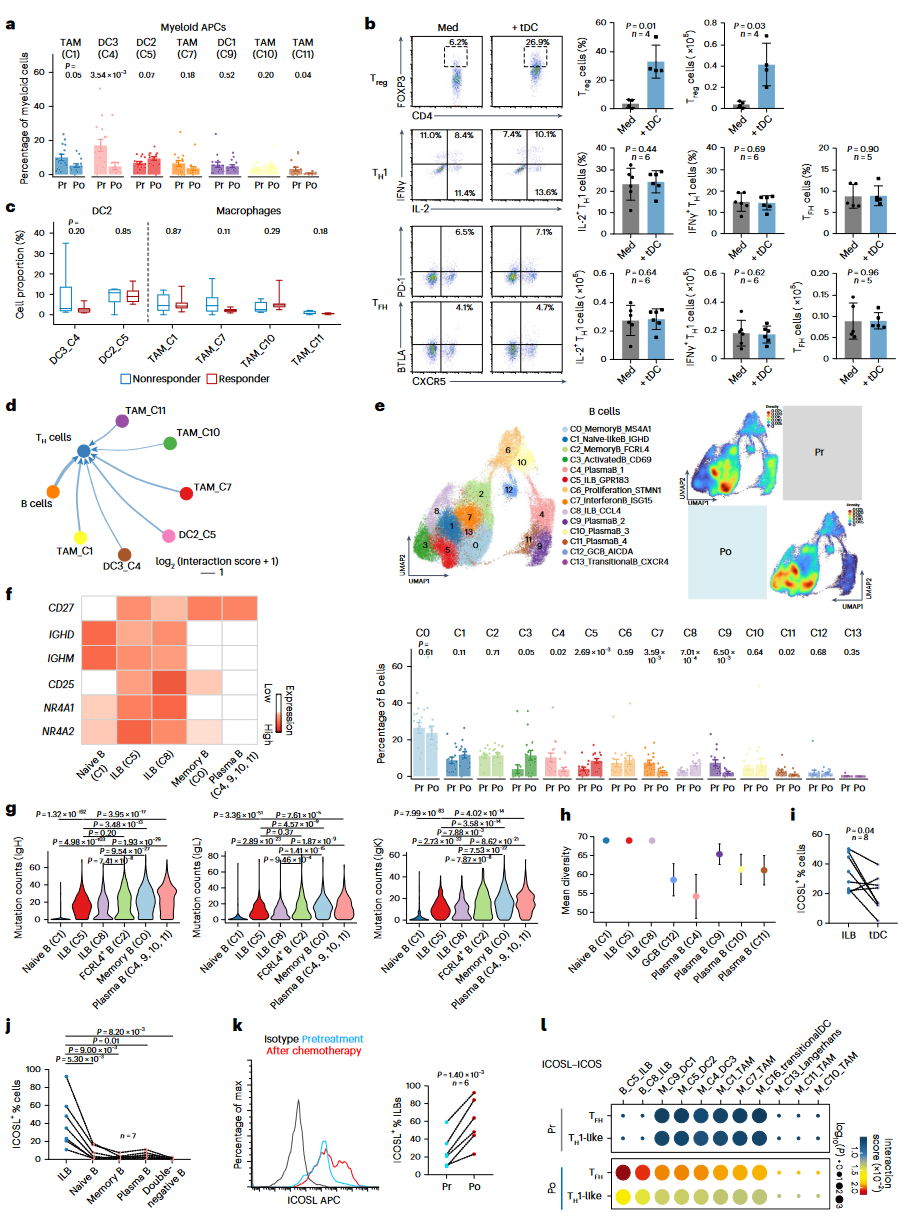
图3:ILB主导的抗肿瘤T细胞免疫的识别
为了进一步探索化疗后ILBs对TIME的贡献,研究团队对化疗后的NPC样本中的ILBs和肿瘤浸润T细胞进行了时空分析。化疗后,在应答者中观察到ILBs显著聚集在类似三级淋巴器官(TLO)的结构中,这些区域也有大量CD4+ T细胞浸润(图4a)。通过对纵向样本(包括未经治疗、化疗后1周、2-3周、4-5周)的详细分析,发现ILBs的频率在化疗开始1周后在TLO样结构中显著增加,而在非应答者中,ILBs的增加非常微弱(图4b)。
进一步分析发现,位于ILBs附近的T细胞增殖水平显著上升,并且增殖T细胞的密度与ILBs呈正相关,但与非ILB的原始B细胞无关(图4c)。在这些区域中,还检测到更多的Th1、Tfh 和GZMB+CD8+ T细胞,而Treg和HAVCR2+CD8+ T细胞较少(图4d)。值得注意的是,化疗后样本中TLO样结构的数量显著增加(化疗前后24名患者匹配样本中分别鉴定出19个TLO和98个TLO样结构;图4e),但化疗后这些结构中没有发现生发中心(GC)区域(图4f)。相反,未经治疗的样本中几乎50%的TLO包含GC(图4f)。更重要的是,这些缺乏GC的TLO样结构在应答者中显著增加(图4g),这一发现通过对纵向肿瘤样本的进一步验证得到支持(图4b)。
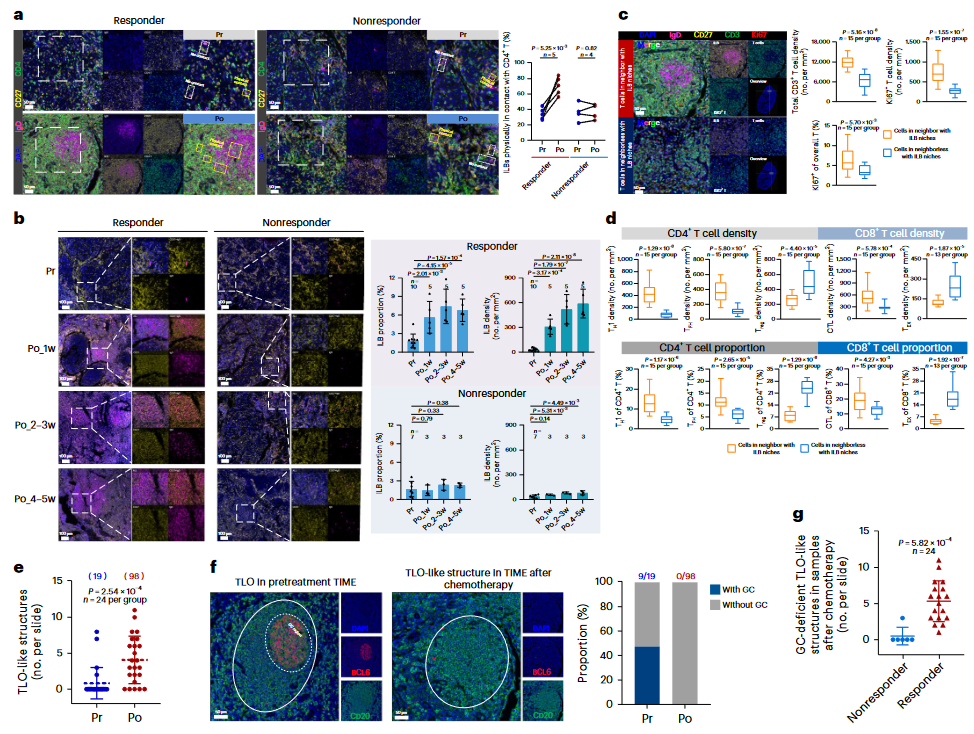
图4:化疗后ILB与Tfh和Th1聚集成TLO样结构
5、ILB通过ICOSL-ICOS轴扩增Tfh和Th1诱导CTL反应
为了探讨ILBs的产生和作用机制,研究采用扩散图谱重构了B细胞的轨迹,结果显示ILBs在化疗过程中从原始B细胞逐步激活并保持稳定轨迹(图5a)。考虑到化疗诱导的DNA片段可以增强癌细胞的MHC I类分子表达,研究进一步探讨了这些DNA片段在诱导ILBs中的潜在作用。实验结果表明,将B细胞与GP化疗处理后的癌细胞培养上清液(GP-SN)共同孵育,可以成功诱导ILBs产生(图5b)。当GP-SN预先用DNase(DNA酶)处理以消耗DNA或阻断B细胞上的TLR9时,ILBs的产生被抑制(图5b),与此一致,TLR9信号及其下游通路在ILBs中显著上调(图5c)。
进一步研究发现,ILBs可以促进Th1和Tfh细胞反应。由于人和小鼠的鼻咽解剖和组织学差异,目前尚无有效的动物模型,因此研究提取从化疗后的NPC患者肿瘤中的肿瘤来源的ILBs和T细胞来进行ILBs功能的体外研究。在共培养系统中,ILBs显示出比非ILB的B细胞更强的扩增IFNγ+IL-2+ Th1和CXCR5+ Tfh细胞的潜力(图5d,e)。值得注意的是,这些ILB扩增的CD4+T细胞在肿瘤裂解物的存在下,增强了CD8+ T细胞表达GZMB和穿孔素的能力,这一过程可以通过中和IL-21来减弱(图5f)。
为了阐明ILB介导的Th1和Tfh扩增机制,研究首先使用特异性抗体屏蔽ILBs中的HLA-DR信号,发现此处理对ILBs诱导的Th1和Tfh扩增几乎没有影响,表明ILBs以MHC II类分子非依赖的方式调节Tfh和Th1细胞(图5g,h)。随后,研究通过阻断T细胞上ICOS受体的信号,发现这种处理有效抑制了ILBs诱导的Th1和Tfh扩增(图5g,h)。ICOS信号通过mTOR依赖的途径触发糖酵解并促进效应T细胞增殖。与此一致,向肿瘤来源的CD4+ T细胞共培养体系中添加ICOS激动剂,显著激活了PI3K–Akt–mTOR通路,而未激活其他信号(图5i)。抑制CD4+ T细胞中的mTOR活性显著减弱了ILBs介导的Th1和Tfh扩增(图5j),这些数据揭示了化疗通过TLR9信号诱导ILBs产生,并通过ICOSL共刺激途径触发Th1和Tfh细胞扩增。
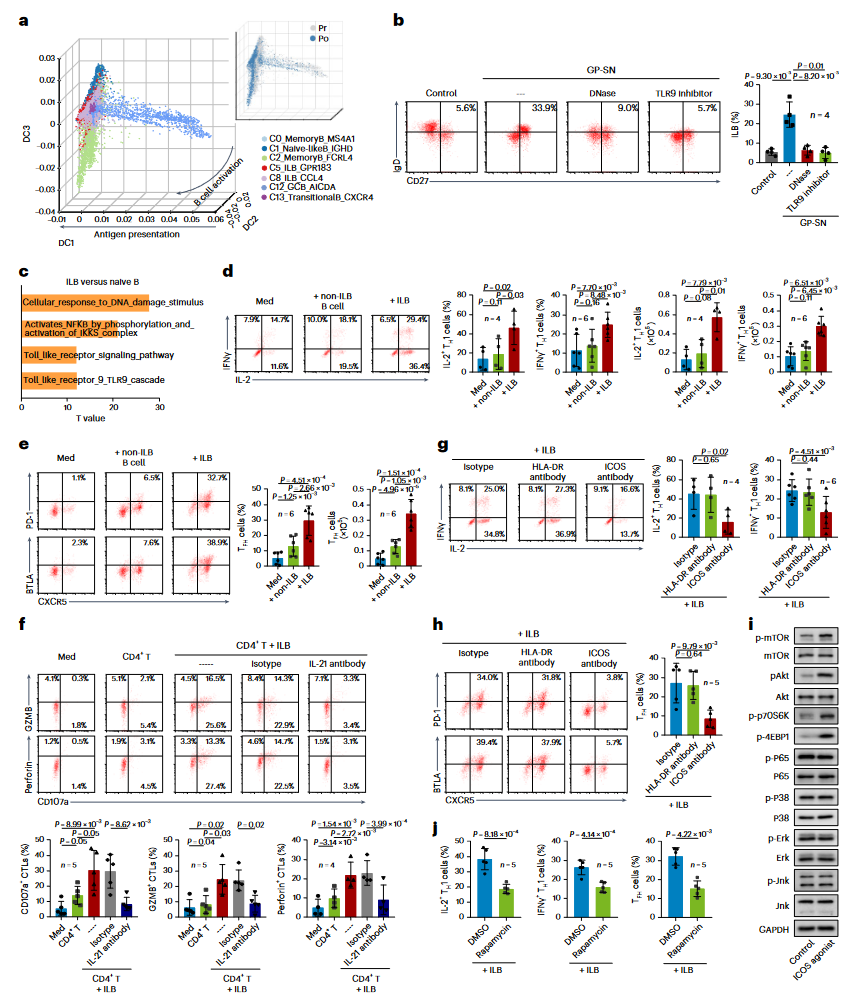
图5:ILB通过ICOSL-ICOS轴扩增Tfh和Th1诱导CTL反应
6、ILBs 预测了鼻咽癌患者对GP方案治疗的疗效
为了探讨ILBs在临床中的意义,研究首先在SYSUCC_E队列(Cohort_1,n = 15对)中分析了化疗后样本中的ILB频率。结果表明,ILB频率能够有效区分化疗反应者和非反应者(反应者为18.5%,非反应者为7.0%,P = 0.04;图6a)。此外,ILBs预测化疗效果的能力优于髓系DC和巨噬细胞(ILBs的曲线下面积AUC = 0.82,DC2的AUC = 0.58,TAM的AUC = 0.52;图6b)。与机制性发现一致,ILB频率与Tfh、Th1和活化T细胞的频率呈正相关,而与Treg细胞无关(图6c)。高ICOSL表达的ILB群体显示出最好的化疗反应效果,相比之下,ILB频率较低或ILBs中ICOSL低表达的患者化疗反应较差(图6d)。重要的是,利用已发表的多中心III期临床试验(NCT01872962)中的患者样本(Cohort_3,GP试验队列,n = 139),研究发现肿瘤浸润ILB频率的增加显著与所有接受GP化疗的NPC患者的无病生存期(DFS)和总生存期(OS)相关(图6e)。此外,ILB高浸润的患者不仅表现出更好的局部肿瘤控制率,还表现出较低的远处转移率(图6e)。多变量Cox回归分析包括已知的预后因素,显示ILBs是独立的良好预后预测因子。进一步通过单细胞数据集计算ILB特征基因集,证实ILB特征也能在SYSUCC_M队列(Cohort_2,n = 150)中预测良好的生存结局(图6f)。
鉴于本研究揭示的GP化疗的免疫调节作用以及GP和免疫检查点抑制剂(ICI)联合治疗在NPC中的临床疗效,研究最后在GP-ICI队列(Cohort_4,n = 380)中测试了ILBs作为GP-ICI联合治疗反应潜在生物标志物的可能性。研究观察到肿瘤浸润ILB频率的增加与确诊的客观反应率显著相关(图6g)。一致地,ILB高浸润的患者具有更长的无进展生存期(PFS)和OS(图6h)。多变量Cox回归分析再次确认ILBs是独立的良好生存预后预测因子(补充表10)。
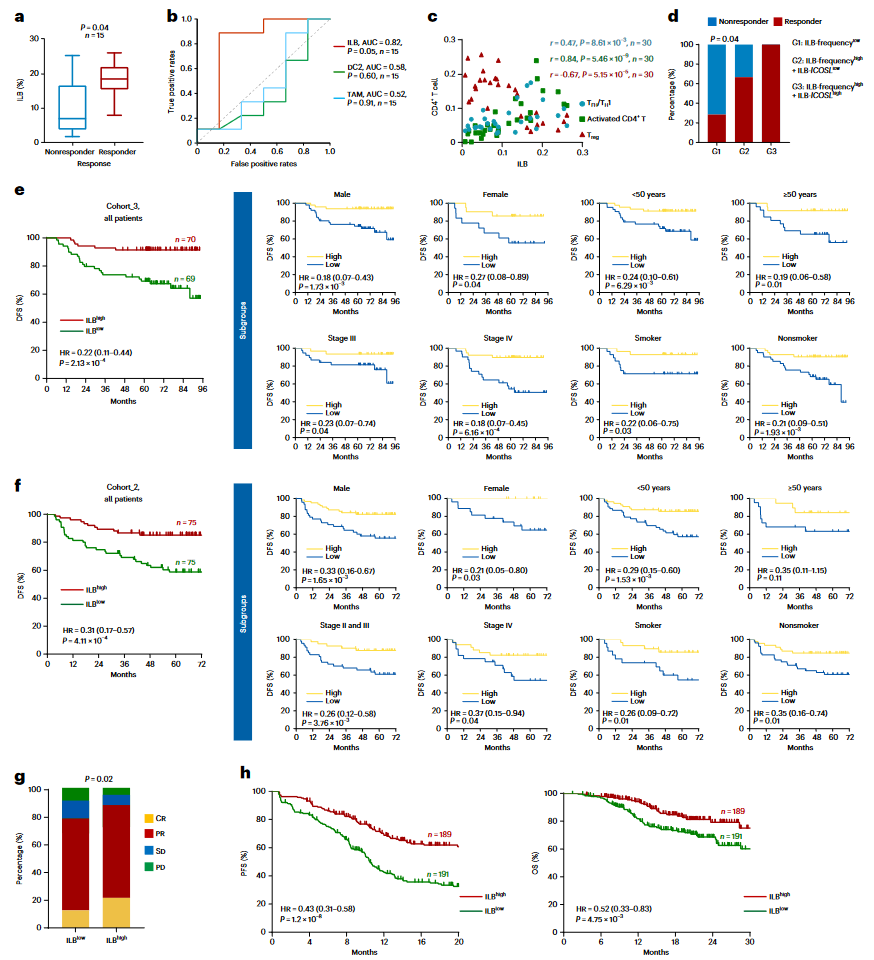
图6:ILBs 预测了鼻咽癌患者对GP方案治疗的疗效
总结:
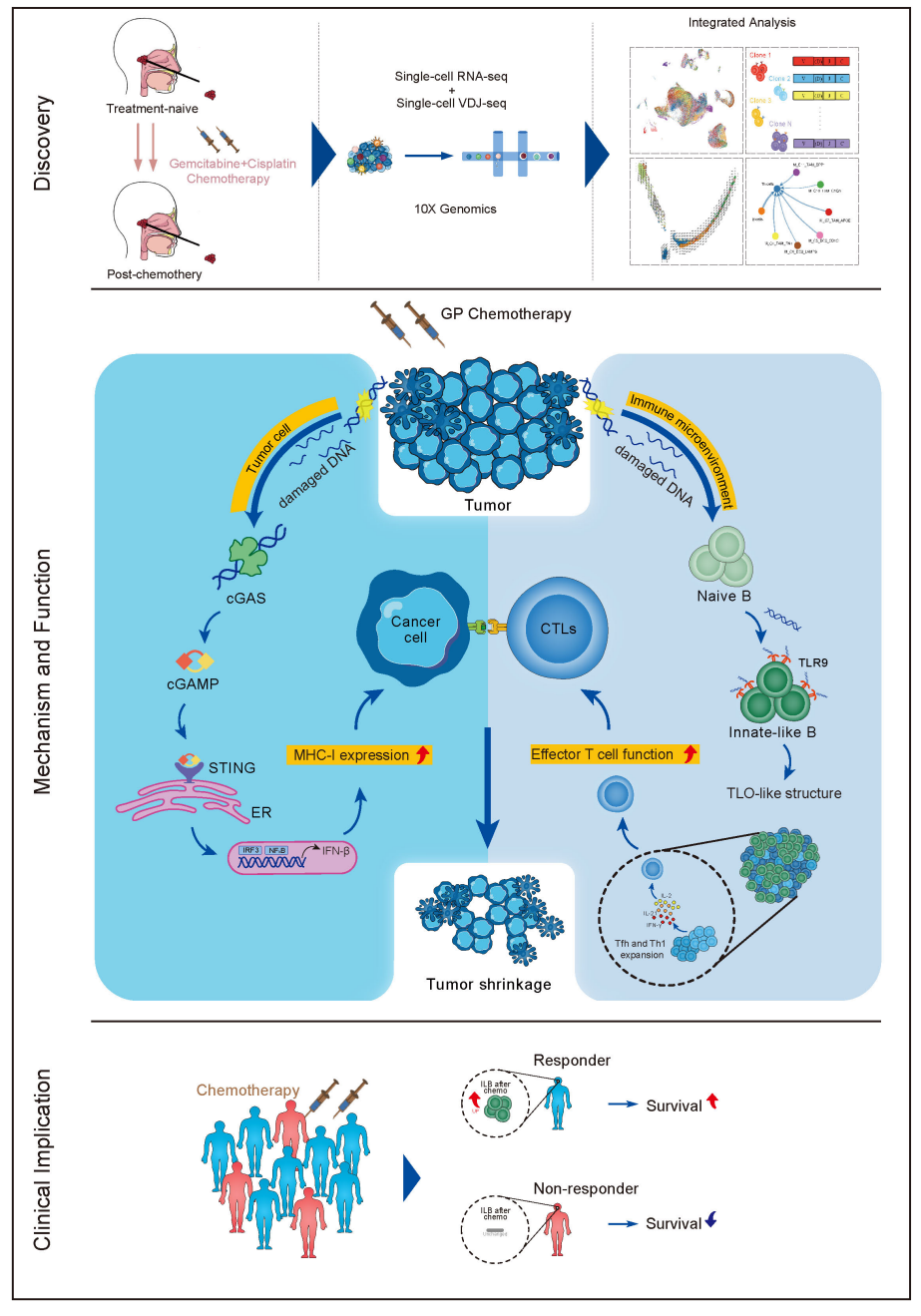
该研究发现,GP化疗介导肿瘤细胞释放DNA片段,这些片段能通过TLR9信号途径诱导一类新识别的细胞亚群——定位于类三级淋巴样结构中的CD27+IgD+IgM+先天样B细胞(ILBs)的产生。同时,这一过程通过ICOSL-ICOS信号轴促进了I型辅助性T细胞(Th1)和滤泡辅助性T细胞(Tfh)的增殖,并增强了细胞毒性CD8+T细胞的杀伤能力。此外,GP化疗还激活了肿瘤细胞内的STING-type-I-interferon通路,上调了肿瘤细胞表面主要组织相容性复合体MHC-I的表达,二者协同作用,形成了一个有效的免疫激活环路,促进了对鼻咽癌细胞的识别和杀伤。
此研究揭示了GP化疗在调控鼻咽癌免疫微环境方面的新机制,明确了肿瘤微环境中存在一种新的ILBs细胞亚群,并验证了这一细胞亚群可作为预测GP化疗效果的生物标志物。该研究还提出了以B细胞为中心的抗肿瘤免疫网络新假说,为鼻咽癌联合治疗新策略的开发提供了重要的理论支撑。

汇报人:毛敏姿
导师:赵宇
审核:程丹妮、任建君
